SSR Mapping for Locus Conferring on the Triple-Spikelet Trait of the Tibetan Triple-spikelet Wheat (Triticum aestivum L. concv. tripletum) 
2. Institute of Plant Protection, Sichuan Academy of Agricultural Sciences, Chengdu, 610066, P.R. China
 Author
Author  Correspondence author
Correspondence author
Triticeae Genomics and Genetics, 2011, Vol. 2, No. 1 doi: 10.5376/tgg.2011.02.0001
Received: 22 Oct., 2011 Accepted: 01 Nov., 2011 Published: 29 Jan., 2012
Li et al., 2011, SSR Mapping Locus Conferring on the Triple-Spikelet Trait of the Tibetan Triple-spikelet Wheat (Triticum aestivum L. concv. tripletum), Triticeae Genomics and Genetics, Vol.2, No.1 1-6 (doi: 10.5376/tgg.2011.02.0001)
Tibetan triple-spikelet wheat is an unique common wheat landrace in Tibet region of China, which has special triple spikelet trait with supernumerary spikelets and florets. Molecular Mapping the control gene locus conferring on triple spikelet trait and mining the closely linked markers would be facilitate high-yield wheat breeding by marker-assisted selection approach. In this study, derived lines TTSW-5 from Tibetan triple spikelet wheat and common-spikelet wheat, Jian 3 and Chuanmai 55, were used to construct the F2 populations for , phenotypic analysis and SSR genotyping. Genetic analysis of phenotypic traits showed that triple spikelet trait of Tibetan triple spikelet wheat are controlled by two independent recessive gene loci. One QTL linked to the triple spikelet trait was detected on the chromosome 2A by using F2 population from TTSW-5/Jian 3 combination and SSR markers, the targeted locus was located within SSR markers, Xgwm275 and Xgwm122, the genetic distance between two markers is 6.6CM with LOD value 6.19, which can be explained 33.1% phenotypic variation, The detected locus tentatively named as qTS2A-1. We speculated that qTS2A-1 locus might be one of dominant loci for controlling the triple spikelet trait , Therefore, SSR markers, Xgwm275 and Xgwm122, might be used as assisted selection markers for triple spikelet trait in high-yield breeding program.
Wheat is the largest cultivated and the most consumed cereal crops in the world to feed more than 35% of the global population as well as to provide the necessary nutrients of human beings in 20% of energy and protein (Sun et al., 2009; Yang et al., 2004). As the development of the world’s economy and changes in the structures of foods in developing countries, particular in the rapid growth of the world's population, wheat is considered to be the crop with the world's largest growth in demand (Rajaram, 2002). It is estimated that global demand for wheat in 2020 will reach 840 million tons to 1050 million tons, which means that wheat production needs to increase by 2.0% annually on the basis of the existing production of 560 million tons (Gill et al., 2004; Sun et al., 2009). Clearly, a substantial increase of wheat production would be the major solution to solve this problem. Therefore, breaking yield barriers and enhancing yields per unit would be the great challenges for wheat breeders. However, breeders applied a small number of well-known backbones of parents in long-term of wheat breeding and improvement, leading to narrow the genetic base of cultivated wheat varieties narrow. There are serious genetic erosion resulting in pandemics of pest and disease as well as great loss of yield in wheat production (Porceddu et al., 1988; Yang et al., 2004), which seriously constrains the further improvements in wheat breeding program. In wheat germplasm resources, the multiple-spikelet wheat (the numbers of spikelet is 30 or more) is specific germplasm with the traits of multiple spikelet and grains having the potential to increase wheat yield. Therefore, Utilization of this specific resource, a kind of multi-spikelet wheat, to change the panicle morphological characters as well as to enhance the number of spikelet and grains have become one of the ideal ways in high-yield breeding of wheat, of which has made encouraging progress so far (Rajaram, 2002; Yen et al., 1995; Swaminathan et al., 1966; Koric, 1973; 1980).
Multiple spikelet traits can be divided into three types, including branch type multiple spikelet, complex spike type multiple spikelet and straight-spike type multiple spikelet (no branch) (Millet, 1987; Li and Zhao, 2000). Tibetan triple spikelet wheat (Triticum aestivum L. Concv. Tripletum, 2n=42, AABBDD), belonging to the straight-spike type multiple spikelet, was a unique landrace collected in 1990’s in the Tibet of China, with the traits of three spikes bearing on each rachis of ear (Figure 1), while common wheat (Triticum aestivum L., 2n=42, AABBDD) has only one spike on each rachis of panicle (Figure 1) (Yan and Yang, 1999; Yang et al., 2004). The characteristics of triple spikelet in Tibetan triple spikelet wheat have genetic stability, similar to the triple spikelet of the six-rowed barley. Although the ear length of Tibetan triple spikelet wheat is quite similar with the ordinary ear length, the plant can bear more than 60 spikes each ear with more than 170 spikelets as well as has the normal tillering and full filling seed, of which the numbers of spikes and spikelets reach 2 or 3 times than that of common wheat (Yang et al., 2004). So far Tibetan triple spikelet wheat is the only one of triple spikelet wheat found in the world, which will provide a new valuable gene resource to mine new favorable genes as well as to develop high yield variety of wheat. Currently, most of studies showed that the trait if multiple spikelet (supernumerary spikelet) would be a recessive trait, governed by one or two genes (Yang et al., 2005; Martinek and Bednar, 2001; Peng et al., 1998a; Millet, 1986; Pennell and Halloran, 1983; Sharman, 1967). Some of studies found that the multiple spikelet might be dominant or partially dominant trait inheritance controlled by one or two genes (Sun et al., 2009; Sun et al., 2000; Chapman and McNeal, 1971; Dencic, 1988). Also some studies found that multiple spikelet traits could be quantitative inheritance, usually jointly controlled by a major gene and several minor genes (Zheng, 1994a; 1994b; Klindworth et al., 1990a; 1990b; Millet, 1987). Previous studies on chromosome analysis by using monomer and nullisome of Chinese Spring had some multiple spikelet gene mapped on the chromosomes of 2A, 4A, 5A, 7A, 2B, 4B, 2D, 3D and 6D (Peng et al., 1998a; 1998b; Zheng et al., 1994b; Yen et al., 1995), of which there was a strong genetic effect on the chromosome 2D (Peng et al., 1998a). It has reported that the second homology group of wheat chromosome might carry the genes controlling trait of multiple spikelet by studies on multiple spikelet characteristics of the tetraploid and hexaploid wheat (Peng et al., 1998a; Laykova et al., 2005; Klindworth et al., 1997;). Sears (1954) found that the deletions in the chromosome 2A or 2D of hexaploid wheat might generate LSS trait of which the gene inhibiting this trait located on chromosome 2DS and 2AL.
|
|
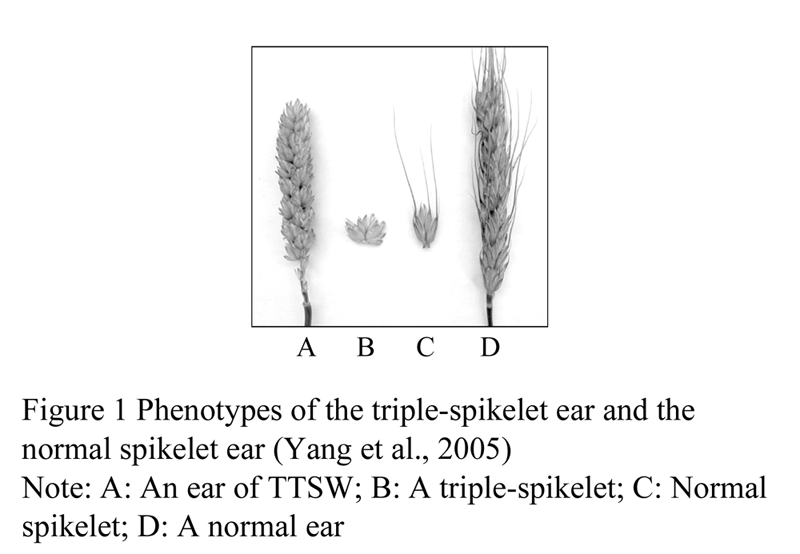
Previous studies of our group have shown that (Yang et al., 2005; Yang et al., 2004) the triple spikelet trait was no any association with the undesirable traits of Tibetan triple spikelet having low kilo-grain weight and low seed setting rate and so on. We obtained excellent triple spikelet individual from the progenies derived the triple spikelet germplasm, which possessed multiple spikelet with more flowers and more grains as well as high weight of 1000 grain weight and single panicle. It has confirmed that it would be feasible to improve the yield of wheat by the use of triple spikelet trait, as well as found that Tibetan triple spikelet trait was controlled by two independent recessive genes based on genetic analysis (Yang et al., 2005; Yang et al., 2004). However, there is no any study on mapping on the the triple spikelet trait so far. In this study, we employed the advanced line TTSW-5 with triple spikelet trait derived from the progenies of the cross between Tibetan triple spikelet wheat and Sichuan common wheat to mate with cultivated common wheat Jian 3 and Chuanmai 55 building F2 populations in order to analyze the genetic characteristics of triple spikelet trait and detect the QTL associated with the triple spikelet trait through SSR markers.
1 Results and Analysis
1.1 Genetic analysis of triple spikelet trait
TTSW-5 was mated with Jian 3 and Chuanmai 55 to generate hybrid F1 plants of 47 and 11, respectively. All of F1 plants self-crossing were to generate F2 plants of 1107 and 176, respectively. The panicle phenotypes of F1 and F2 were identified in ear maturity. The results showed that all of F1 from the crosses of TTSW-5/Jian 3 and TTSW-5/Chuanmai 55 exhibit the ear phenotypes of common wheat as the same as the parents of Jian 3 and Chuanmai 55, indicating that trait of triple-spikelet of TTSW-5 would be controlled by a recessive gene.
F2 individuals derived from TTSW-5/Jian 3 and TTSW-5/Chuanmai 55 were phenotyped in mature ear, in which 1040 of 1107 individuals of F2 population from TTSW-5/Jian 3 exhibited the same ear phenotype as the parental Jian 3, while 67 individuals exhibited the triple-spikelet-like trait in different levels from 30%~100% of triple–spikelet panicles accounting for the total spikelets. Likewise, 161 of 176 individuals of F2 population from TTSW-5/Chuangmai 55 exhibited the same ear phenotype as the parental Chuanmai 55, while 15 individuals exhibited the triple-spikelet-like trait in different levels from 35% to 100% of triple–spikelet panicles accounting for the total spikelets. The chi-square test showed that the ratios of two F2 populations were χ2<χ20.05 (χ20.05=3.841), being consistent to 15 to 1 (Table 1), which indicated that triple spikelet traits of TTSW-5 should be governed by two independent genetic recessive genes, this result was identical to our previous results in genetic analysis of population derived from Tibet triple spikelet wheat itself.
|
|
1.2 Polymorphic analysis among parents and triple spikelet and common spikelet
944 SSR markers were chosen from the genomes of A, B and D in common wheat to screen the polymorphisms between the triple spikelet wheat TTSW-5 and Jian 3 of cultivated common wheat, of which 189 markers were detected to be differences accounting for 20% of the total markers with well electrophoresis. 69 polymorphic markers were detected in A genome with 7.3% of polymorphic ratio, 66 polymorphic markers detected in B genome, with 7.0% of polymorphic ratio and 54 polymorphic markers detected in D genome, 5.7% of polymorphic ratio. 14 of 189 SSR markers exhibiting polymorphisms between the parents, including Xgwm377, Xgwm296, Xcfd160, Xcfa2263, Xwmc144, Xgwm95, Xbarc1064, Xbarc11, Xbarc15, Xgwm122, Xgwm425, Xgwm275, Xgwm296 and Xwmc453, showed polymorphisms between the pools in the triple spikelet and the common spikelet wheat.
1.3 QTL detection
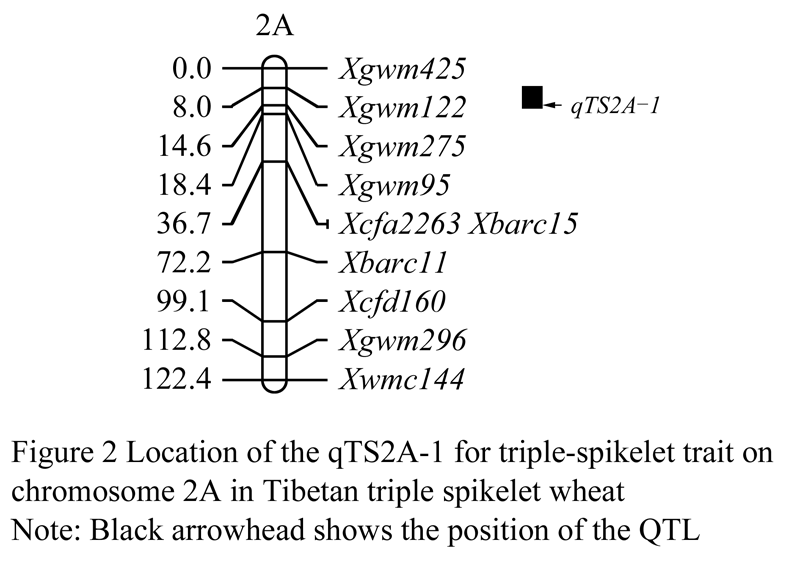
The markers having ploymorphisms in the pools and parents of the triple spikelet and common spikelet wheat were applied to scan the F2 population derived from TTSW-5/Jian 3 for the separation of the triple spikelet trait. Genetic linkage map of chromosome 2A were built by 10 SSR markers covering the 112.8 cM in length of total chromosome (Figure 2). A QTL was detected in 2A on chromosome 1 by composite interval mapping analysis with LOD value of 6.19, tentatively named qTS2A-1, explaining 33.1% of the phenotypic variation. The detected QTL was located between SSR markers, Xgwm275 and Xgwm122, in span of 6.6 cM in the genetic distance (Figure 1). The results indicated that qTS2A-1 might be the major QTL associated with the triple spikelet traits. However, previous genetic analysis showed that the triple spikelet trait of TTSW-5 might be controlled by two independent genetic recessive genes, but there was only a major QTL for triple spikelet trait detected in this study, indicating that another one should not detected yet. It might be possible due to limited SSR markers or too far distance among SSR markers. So regarding another recessive locus, we managed to us new genetic population to identify the location and its effect of the tentative locus.
|
|
2 Discussions
A lot of analysis on genetic models of multiple spike traits has been made already. Patil (1958) considered that the branching traits of durum wheat might be controlled by single recessive gene. Pennell and Halloran (1983) studied that unstable branching trait of durum wheat lines AU S4531 controlled by two recessive genes. Aybeniz et al (2011) found that the novel branching panicle traits derived from the hybrid of synthesized wheat line 171ACS and hardy wheat were also controlled by the single recessive gene. Sun et al (2009) studies have shown that multiple spikelet wheat line 51885 was controlled by two pairs of dominant genes. Koric et al (1996) study suggested that branching panicle traits of common wheat were controlled by two pairs of dominant complementary and pyramiding genes. Our research group has used the Tibetan triple spikelet wheat to mate with Chuanmai 38 and SW8488, respectively to generate F2 and backcross populations for genetic analysis, indicating triple spikelet trait of Tibetan triple spikelet was controlled by two independent recessive genes (Yang et al., 2005; Yang et al., 2004). Although the genetic model of multiple spikelet traits might be difference due to using different materials that may carry different genes, our results in this study had confirmed the above mentioned conclusions previous done by our research group (Yang et al., 2005; Yang et al., 2004).
Klindworth et al (1990b) found a recessive major gene associated with branching trait in the chromosome 2AS, but its linkage molecular marker was unavailable. Qxana et al (2009) had multiple spikelet gene Mrs1 mapped on chromosome 2DS, co-separating with SSR locus Xwmc453, meanwhile rye multiple spikelet gene Mol gene mapped on chromosome 2R. Although in this study the maim effective QTL closely related to triple spikelet traits was detected on chromosome 2A, a multiple spikelet locus detected by Qxana, Xwmc453, using to scan the parents and populations did not exhibit any correlation with the triple spikelet trait of Tibetan triple spikelet wheat, indicating that the QTL detected in this study should be difference from the Mrs1 locus mapped by Qxana et al (2009).
3 Materials and Methods
3.1 Materials used in this study
Tibetan triple spikelet wheat is a unique landrace in Tibet of China provided by the Sichuan Agricultural University, possessing similar triple spikelet trait of six-rowed barley with more than 60 spiklets and more than 170 florets, which should be 2 or 3 times than that of common wheat. TTSW-5 is the triple spikelet line derived from the progenies of the cross between the Tibetan triple spikelet wheat and Sichuan wheat Chuanmai 38. Jian 3 is the breeding line of common wheat developed by Crops Research Institute of Sichuan Academy of Agricultural Sciences. Chuanmai 55 is new variety of common wheat developed by Sichuan Academy of Agricultural Sciences. F2 populations from the cross between triple spikelet wheat TTSW-5 and Jian 3 was employed for triple spikelet trait mapping and QTL detection. F2 population from the cross of TTSW-5 and Chuanmai 55 were used for further genetic validation on phenotype.
3.2 Phenotypic identification
TTSW-5 was mated with Jian 3 and Chuanmai 55 to generate hybrid F1 plants of 47 and 11, respectively. All of F1 plants self-crossing were to generate F2 plants of 1107 and 176, respectively. The panicle phenotypes of F1 and F2 were identified in ear maturity. The phenotypic methods were as following: harvesting the mature individual, measuring the number of triple spikelet per ear and total number of spikelet of whole ear to calculate the percentage by the number of triple spikelet to total of spikelet, the average percentage of each plant was assigned to be a phenotypic data of each ear.
3.3 DNA extraction and SSR analysis
Genomic DNA was extracted from the parents and F2 individuals derived from TTSW-5/Jian 3 by using CTAB method. 944 SSR markers encoded with Barc, Xgwm, Xgdm, Xwmc, Xcfd etc. (Röder et al., 1998; Pestsova et al., 2000; Somers et al., 2004; Song et al., 2005) applied to scan the polymorphism between TTSW-5 and Jian 3. DNA pool of triple spikelet and DNA pool of common spikelet were established by using eight triple spikelet individuals and the common according to the results of phenotypic identification, respectively, find the polymorphism locus between the traits of triple spikelet and common spikelet by using the bulked segregant analysis (BSA) method.
PCR reaction was performed in the total volume of 20 μL containing 1×buffer (100 mmol/L Tris-HCl, pH 8.3, 1.5 mmol/L MgCl2), 0.2 mmol/L dNTPs, 50 ng random primers, 1 U Taq DNA polymer enzyme, 50~100 ng template DNA. PCR amplification procedures were following as 94℃ pre-denaturing for 5 min and then following 35 cycles as 94℃ denaturing for 1 min, 50℃/55℃/60℃ annealing for 1 min, 72℃ extending for 1 min and finally 72℃ extending for 10 min. PCR amplification was carried out in a PTC-200, SSR primers were synthesized by the TaKaRa Company. PCR products was scored by 6% polyacrylamide gel electrophoresis with 1×TBE of electrophoresis buffer and 400 V voltages for 40 min, stained with nitrate silver and digitalized by digital camera.
3.4 Genetic linkage map and QTL analysis
Linkage map was constructed by the Software of Map Manager QTXb20, recombination rate was converted to genetic distance (centiMorgan, cM) with Kosambi function. QTL were detected by using composite interval mapping of QTL Cartographer 2.5 software.
Author’s contribution
JL completed the experimental design, result analysis and manuscript preparation and revision; JW, HTW and XRH are the main executor of the experimental work, completed data analysis and involved in writing. WYY conceived the project and designed the experiments as well as wrote and revised manuscript. All authors had read and agreed the final text.
Acknowledgement
This study was jointly supported by the National Foundation of Natural Sciences (30871532), National Key Basic Research Projects (2011CB100100), Specific Funds of Modern Agricultural Technology System (CARS-3-2-41), Sichuan Provincial Breeding Project, Sichuan International Cooperation Project (2010HH0052, 2011HH0026) and Youth Fund and Excellent Breeding Articles of Sichuan Province Finance.
References
Aybeniz J.A., and Naib K.A., 2011, Inheritance of the branching in hybrid populations among tetraploid wheat species and the new branched spike line 166-Schakheli, Genet. Resour. Crop Ev., 58: 621-628
http://dx.doi.org/10.1007/s10722-011-9702-9
Benito C., Zaragoza C., Gallego F.J., De la Pena A., Figueiras A.M., 1991, A map of rye chromosome 2R using isozyme and morphological markers, Theor. Appl. Genet., 82: 112-116
http://dx.doi.org/10.1007/BF00231284
Chapman S.R., and McNeal F.H., 1971, Gene action for yields components and plant height in spring wheat cross, Crop Sci., 11: 384-386
http://dx.doi.org/10.2135/cropsci1971.0011183X001100030022x
De Vries J.N., and Sybenga J., 1984, Chromosomal location of 17 monogenically inherited morphological markers in rye (Secale cereale L.) using the translocation tester set, Zeitschrift für Pflanzenzüchtung, 92: 117-139
Dencic S., 1988, Genetic analysis of different structures of sink capacity in wheat, In: Miller T.E., Koebner R.M.D, (eds.), Proc. 7th Int., Wheat Genet. Symp., Institute of Plant Science Research, Cambridge, UK, pp.499-502
Gill B.S., Appels R., Botha-Oberholster A.M., Buell C.R., Bennetzen J.L., Chalhoub B., Chumley F., Dvorák J., Iwanaga M., Keller B., Li W.L., McCombie W.R., Ogihara Y., Quetier F., and Sasaki T., 2004, A workshop report on wheat genome sequencing, International Genome Research Wheat Consortium, Genetics, 168: 1087-1096
http://dx.doi.org/10.1534/genetics.104.034769 PMid:15514080 PMCid:1448818
Klindworth D.L., Williams N.D., and Joppa L.R., 1990a, Inheritance of supernumerary spikelets in a tetraploid wheat cross, Genome, 33: 509-514
http://dx.doi.org/10.1139/g90-075
Klindworth D.L., Williams N.D., and Joppa L.R., 1990b, Chromosomal location of genes for supernumerary spikelets in tetraploid wheat, Genome, 33: 515-520
http://dx.doi.org/10.1139/g90-076
Klindworth D.L., Klindworth M.M., and Williams N.D., 1997, Telosomic mapping of four genetic markers in durum wheat, J. Hered., 88: 229-232
Koric S., 1973, Branching genes in Triticum aestivum, In: Sears E.R., Sears L.M.S.(eds.), Proceeding of the 4th international wheat genetics symposium, Columbia, Mo., USA, pp.283-288
Koric S., 1996, Productivity of branched ears of vuglare wheat, Savrem. Poljopr., 14: 545-552
Koric S., 1980, Study of branched gene complex of Triticum aestivum ssp. vulgare and its significance for wheat breeding, J. Sci. Agric. Res. Zagreb. Yugoslavia, 142: 271-282
Laykova L.I., Arbuzova V.S., Popova O.M., Efremova T.T., and Melnick V.M., 2005, Study on spike branching in the T. aestivum mutant lines (cv. Saratovskaya29). In: Goncharov P.L., Zilke R.A., and Gordeeva T.N. (eds.), Proceeding of the IX workshop on genetics and breeding, Novosibirsk, Russia, pp.388-393
Li W.P., and Zhao W.M., 2000, A breeding method for increasing spikelet and studies on creation of new germplasm resource in wheat, Zuowu Xuebao (Acta. Agron. Sin.), 26(2): 222-230
Li J., Wei H.T., Hu X.R., Li C.S., Tai Y.L., Liu D.C., Yang W.Y., 2011, Identification of a high-yield introgression locus from synthetic hexaploid wheat in Chuanmai42, Zuowu Xuebao (Acta. Agron. Sin.), 37(2): 255-262
http://dx.doi.org/10.1016/S1875-2780(11)60007-2http://dx.doi.org/10.3724/SP.J.1006.2011.00255
Millet E., 1986, Genetic control of heading date and spikelet number in common wheat (Triticum aestivum L.) Line ‘Noa’, Theor. Appl. Genet., 72:105-107
Millet E., 1987, Monosomic analysis of heading date and spikelet number in common wheat (Triticum aestivum L.) multispikelet line Noa, Theor. Appl. Genet., 74: 487- 492
http://dx.doi.org/10.1007/BF00289827
Martinek P., and Bednar J., 2001, Changes of spike morphology (multirow spike—MRS, long glumes—LG) in wheat (Triticum aestivum L.) and their importance for breeding, In: The proceedings of international conference “genetic collections, isogenic and alloplasmic lines”, Novosibirsk, Russia, pp.192-194
Oxana D., Petr M., Anatoly V.V., Viktor K., Marion S., Röder M.S., and Andreas B., 2009, Microsatellite mapping of genes that determine supernumerary spikelets in wheat (T. aestivum) and rye (S. cereale), Theor. Appl. Genet., 119: 867-874
http://dx.doi.org/10.1007/s00122-009-1095-1 PMid:19568730
Patil J.A., 1958, Inheritance study in wheat, Curr. Sci., 27: 404-405
Pestsova E., Ganal M.W., and Röder M.S., 2000, Isolation and mapping of microsatellite markers specific for the D genome of bread wheat, Genome, 4:689–697
http://dx.doi.org/10.1139/g00-042 PMid:10984182
Peng Z.S., Liu D.C., Yen C., and Yang J.L., 1998a, Genetic control of supernumerary spikelet in common wheat line LYB, Wheat Information Service, 86: 6-12
Peng Z.S., Yen C., and Yang J.L., 1998b, Chromosomal location of genes for supernumerary spikelet in bread wheat, Euphytica, 103:109-114
http://dx.doi.org/10.1023/A:1018323310621http://dx.doi.org/10.1023/A:1018383612439
Pennell A.L., and Halloran G.M., 1983, Inheritance of supernumerary spikelets in wheat, Euphytica, 32: 767-776
http://dx.doi.org/10.1007/BF00042157
Porceddu E., Ceoloni C., Lafiandra D., Tanzarella O.A., and Scarascia Mugnozza G.T., 1988, Genetic resources and plant breeding: problems and prospects, In: Miller T.E., Koebner R.M.D.(eds.), Proc. 7th Intern. Wheat Genet. Symp., Institute of Plant Science Research, Cambridge, UK, pp.7- 21
Röder M.S., Korzun V., Wendehake K., Plaschke J., Tixier M.H., Leroy P., and Ganal M.W., 1998, A microsatellite map of wheat, Genetics, 149: 2007-2023
PMid:9691054 PMCid:1460256
Rajaram S., 2002, Prospects and promise of wheat breeding in the 21st century, In: He Z.H., Zhang A.M. (eds.), Advance of wheat breeding in China, China Science and Technology Press, Beijing, pp.38-53
Sears E.R., 1954, The aneuploids of common wheat, University of Missouri, Columbia, Mo, pp.3-58
Swaminathan M.S., Chopra V.L., and Sastry G.R.K., 1966, Expression and stability of an induced mutation for ear branching in bread wheat, Curr. Sci., 35: 91-92
Sharman B.C., 1967, Interpretation of the morphology of various naturally occurring abnormalities of the inflorescence of wheat (Triticum ssp.), Can. J. Bot., 45: 2073-2080
http://dx.doi.org/10.1139/b67-224
Sun D.F., Fang J.Y., and Sun G.L., 2009, Inheritance of genes controlling supernumerary spikelet in wheat line 51885, Euphytica, 167: 173-179
http://dx.doi.org/10.1007/s10681-008-9854-7
Sun D.F., Zhu X.D., Wan Z.B., and Cai J., 2000, The stability and expression in F1 generation of supernumerary spikelets in bread wheat, Huazhong Nongye Daxue Xuebao (J. Huazhong Agric. Univ.), 19: 213-218
Somers D.J., Isaac P., and Edwards K., 2004, A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.), Theor. Appl. Genet., 109: 1105-1114
http://dx.doi.org/10.1007/s00122-004-1740-7 PMid:15490101
Song Q.J., Shi J.R., Singh S., Fickus E.W., Costa J.M., Lewis J., Gill B.S., Ward R., and Cregan P.B., 2005, Development and mapping of microsatellite (SSR) marker in wheat, Theor. Appl. Genet., 110: 550-560
http://dx.doi.org/10.1007/s00122-004-1871-x PMid:15655666
Yang W.Y, Lu C.M, Lu B.R, Wang Y., Hu X.R., Yu Y., and Zang Y., 2004, Inheritance of the character of triple spikelets in a Tibetan triple spikelet wheat (Triticum aestivum L concv. Tripletum), Xinan Nongye Xuebao (South west China Journal of Agricultural Sciences), 17(1): 1-4
Yang W.Y., Lu B.R., Hu X.R., Yu Y., and Zhang Y., 2005, Inheritance of the triple-spikelet character in a Tibetan landrace of common, Genet. Resour. Crop Ev., 52: 847-851
http://dx.doi.org/10.1007/s10722-003-6089-2
Yen C., Zheng Y.L., and Yang J.L., 1995, An ideotype for high yield breeding, in theory and practice. In: Li Z.S., Xin Z.Y. (eds.), Proc. 8th Intern. Wheat Genet. Symp., Beijing, China, pp.1113- 1117
Yen C. and Yang J.L., eds., 1999, Triticeae Bio-Systematics, Wheat-Aegilops complex, China Agricultural Press, Beijing, China, pp.114-116
Zheng Y.L., Yan C., and Yang J.L., 1994a, Gene effects and their correlation analysis of four specific characters in the common wheat multispikelet line”10-A, Zuowu Xuebao (Acta. Agron. Sin.), 20(5): 536-541
Zheng Y.L., Yan C., and Yang J.L., 1994b, Effects of chromosomes on grain yield per plant in the common wheat multispikelet line10-A, Yichuan (Hered.), 16(4): 27-30



. PDF(197KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Jun Li
. Qin Wang
. Huiting Wei
. Xiaorong Hu
. Wuyun Yang
Related articles
. Tibetan triple-spikelet wheat
. Triticum aestivum L. concv. tripletum
. Tibetan wheat landrace
. Triple-spikelet trait
. SSR mapping
. Wheat high-yield breeding
Tools
. Email to a friend
. Post a comment